Distinction entre les traits qualitatifs et quantitatifs chez les poissons
Dans cet article, nous discuterons de la distinction entre les traits qualitatifs et quantitatifs chez les poissons.
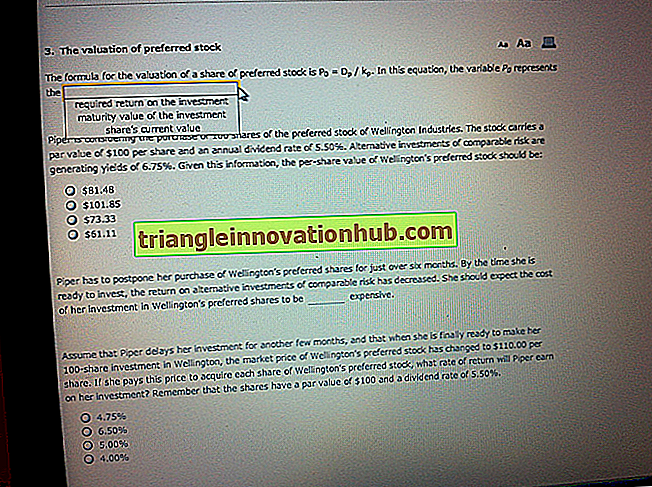
Il existe deux principales catégories phénotypiques de traits (caractères), l’une qualitative et l’autre quantitative. Le schéma de transmission des caractères quantitatifs peut maintenant être largement utilisé en aquaculture pour obtenir des poissons présentant le caractère phénotypique souhaité, en fonction des besoins des animaux marqués.
Le caractère qualitatif peut être distingué phénotypiquement (extérieurement) du caractère quantitatif. Le trait qualitatif est défini en termes qualitatifs, par exemple la couleur des fleurs est rouge ou blanche. Un autre exemple est que la couleur des yeux de Drosophila est soit rouge (sauvage) ou blanche (mutant). Ces couleurs sont des catégories distinctes ou des variantes de couleurs distinctes.
La couleur de la variété sauvage de Cyprinus carpio est noire, mais dans la population, certains poissons présentent une couleur dorée (variante). Les poissons de couleur dorée sont très demandés sur le marché européen. Ces exemples sont des exemples de variations qualitatives ou discontinues.
Les variations quantitatives sont les variations du poids corporel à différents âges, le taux de croissance larvaire et le pourcentage de graisse dans la viande, le poids du poisson, le temps requis pour que le poisson atteigne sa maturité, la taille des nageoires ou la forme des nageoires sont l'exemple des variations quantitatives. . La maturité précoce et la quantité de viande dans le poisson sont importantes pour les aquaculteurs.
Les traits qualitatifs et quantitatifs pourraient également être distingués sur la base de l'héritage en fonction de leur combinaison de génotype spécifique. Si la progéniture de la génération F 1 ressemble phénotypiquement à l’un des parents alors que, dans F 2, le rapport phénotypique de la progéniture indique respectivement 3: 1 (3/4 ou ¼) dominant / récessif.
Si les roses blanches sont croisées avec des roses rouges et que la couleur des roses dans F 1 est soit rouge, soit blanche (ressemble à l’un des parents), c’est le caractère qualitatif. Un autre exemple d'héritage qualitatif est le pelage rouge et blanc chez les bovins.
Si, dans la génération F 1, la couleur phénotypique serait rouge ou blanche et que la couleur phénotypique F 2 serait de 3: 1 dans le croisement monohybride, il est clairement indiqué que le trait de couleur est qualitatif et non quantitatif.
Dans Cyprinus carpio, si les poissons de couleur noire (sauvages et dominants) sont croisés avec des poissons de couleur dorée (variant, récessifs), et dans la génération F 1, tous les poissons sont noirs et si dans la génération F 2, le rapport est de 3: 1 (3 un exemple clair de phénotype qualitatif).
D'autre part, si la couleur phénotypique de la génération F 1 serait intermédiaire entre les couleurs du parent et de la génération F 2, elle serait sous forme de distribution continue entre les couleurs des parents, on parle alors de quantité. héritage (Fig. 40.1).

À l'aide de combinaisons phénotypiques et génotypiques, il est possible de tirer avantage de la production par les pisciculteurs des caractères requis dans le poisson des aquaculteurs. Depuis la redécouverte de la loi de Mendel en 1900, loi de ségrégation et loi d'assortiment indépendant, la controverse a soulevé le fait que la loi de Mendel n'était pas respectée.
Cependant, il est résolu que les traits quantitatifs et qualitatifs suivent l’héritage mendélien. L’incompatibilité est due à la variation des allèles d’un ou de plusieurs locus et aux caractères quantitatifs contrôlés par plusieurs gènes.
Enfin, il est établi que l'hérédité du trait quantitatif dépend de la combinaison du gène de l'allèle, due à des mutations, et que, dans leur totalité, ces traits apparaissent en raison du comportement des gènes et du modèle de ségrégation au cours de la méiose. Il est donc important de connaître le modèle d’héritage.
Le gène est une longueur d'ADN qui contient des milliers ou des centaines de milliers de paires de nucléotides, et les séquences de paires de nucléotides de cet ADN contiennent les informations qui en font le gène.
Selon Mendel, un seul caractère phénotypique d'un organisme est contrôlé par deux allèles ou allélomorphes. On ne savait pas à l'époque que les allèles pouvaient avoir de nombreuses relations alléliques différentes. On sait que les gènes ont non seulement deux allèles, mais également plusieurs allèles.
Nous savons aujourd'hui que les produits de différents gènes interagissent et que ceux des allèles d'un gène génèrent un phénotype différent selon que l'individu possède également des allèles particuliers d'un autre gène. Il est maintenant suggéré que les gènes mendéliens contrôlent aussi bien les traits quantitatifs que qualitatifs.
Les allèles ou allélomorphes d'un même gène interagissent de différentes manières. Il est admis que la mutation a également eu lieu. Ces allèles mutants peuvent donner une large gamme de phénotypes différents. Par exemple, plus de 350 allèles du gène de la fibrose kystique humaine ont été découverts.
Les relations dans le monde biologique sont les suivantes:
Ces relations sont également observées chez les poissons et il est maintenant possible d’obtenir des variantes phénotypiques amicales marquées,
(1) domination complète sur les autres allèles lorsque les allèles sont situés dans les mêmes locus,
(2) Dominance incomplète ou partielle.
(3) Sur domination.
(4) Co-domination (expression égale).
A. Les allèles du même gène-loci peuvent montrer une domination totale:
Les exemples courants donnés par Mendel sont les grands et les nains des pois. La combinaison génétique serait homozygote dominant (TT), hétérozygote (Tt) et homozygote récessif (tt). Selon la loi de Mendel sur l'héritage, les gènes n'ont que deux allèles: l'un dominant et l'autre récessif.
Ces deux gènes sont situés aux mêmes locus, cependant, nous savons maintenant qu’un gène peut avoir plusieurs allèles au même locus. Si l'organisme a hérité de la même version du gène des deux parents, il est dit homozygote dominant / récessif (TT, tt). Si la version est différente de celle qu’elle est hétérozygote (Tt). Chaque version d'un gène particulier s'appelle allèle.
Dans la génération F 1 sur le croisement entre de hautes plantes naines homozygotes et homozygotes, la progéniture aura des plantes hautes, montrant une domination complète sur les plantes naines, mais dans la génération F 2, la progéniture sera à la fois grande et naine dans un rapport de 3: 1. (Croix monohybride ). L’exemple d’une dominance totale des poissons au profit des aquaculteurs est la couleur de corps de Cyprinus carpio.
Dans la nature, la couleur de corps des poissons est grise, mais dans la population, on trouve également des poissons de couleur dorée. Ces poissons de couleur dorée sont appelés comme variantes. Le variant est un allèle ou un phénotype qui diffère du type standard ou sauvage mais n'est pas délétère ou anormal, donc survécu.
La couleur dorée du phénotype qui diffère de la couleur grise standard de type sauvage est due aux mutations génotypiques des allèles que le poisson obtient par héritage. La variante en or a une valeur marchande élevée dans les pays européens. La couleur dorée phénotypique est un trait qualitatif. Les combinaisons génétiques sont connues.
Dans Cyprinus carpio, deux allèles de couleur sauvage sont attribués à un allèle, W et une couleur dorée variant en raison de la présence de l’allèle, G. La couleur dorée du phénotype apparaît dans le poisson si deux allèles homozygotes (GG) existent dans la progéniture à le même locus. Les progénitures ayant la combinaison de génotypes homozygote dominant (WW) ou hétérozygote (WG) montreront une coloration sauvage si elles existent au même locus.
Comme dans ce cas, l'allèle sauvage (W) est dominé tandis que l'allèle d'or (G) est récessif et ces allèles sont présents dans les mêmes locus. Ceci est un exemple simple et parfait de relation de dominance récessive, car le phénotype (couleur dorée) n’est pas apparu à l’état hétérozygote.
Les allèles du même gène peuvent montrer une dominance complète dans une condition homozygote dominante ou hétérozygote. Un phénotype de couleur dorée apparaîtra s'il y a une forme récessive homozygote.
Deux allèles (allélomorphes) d'un même gène montrent une domination complète sur l'action de l'autre qui est récessif. Cela donne les rapports classiques de monohybride, 3: 1 (Fig. 40.2) ou dihybride 9: 3 3: 1. Il s’agit d’une interaction génétique intra-allélique ou allélique. Ce qui suit serait le résultat d'un croisement monohybride et est représenté comme suit (Fig. 40.2).

Profitant de ce fait, l’éleveur doit sélectionner des mâles et des femelles de couleur dorée (récessifs homozygotes) et les reproduire en écloserie pour ne produire que des poissons de couleur dorée. La couleur dorée est rare dans la nature.
B. Les allèles du même gène peuvent montrer une dominance incomplète (semi-dominance):
La dominance peut être incomplète, les allèles semi-dominants peuvent produire le même produit mais en quantité moindre. Un schéma d'héritage de semi-dominance est également observé chez les poissons.
Si la couleur dorée de Cyprinus carpio est l’exemple de la dominance totale, celle des espèces de Tilapia, Oreochromis mossambicus, est le cas évident de la domination incomplète. Le poisson de type sauvage a une couleur noire normale avec une combinaison homozygote comme WW.
La variante a une coloration dorée en tant que GG et la coloration dorée apparaîtra dans le poisson, si le poisson a des allèles homozygotes en tant que GG au même endroit. Si ces poissons sont croisés, la génération F 1 aura un poisson hétérozygote avec la combinaison de génotypes WG. Le poisson hétérozygote ayant la combinaison WG a une couleur bronze au lieu d'être noir ou doré, semblable à celle des parents.
La présence de couleur phénotype bronze dans F 1 indique clairement qu'il n'y a pas de situation de dominance et de récession, mais que la dominance est incomplète (semi-dominance). En semi-dominance, l'hétérozygote montre un phénotype intermédiaire entre le dominant et le récessif.
L’Institut de recherche sur les pêches de Taiwan a mis au point des variantes rouges d’O. Mossambicus contrôlées par des allèles récessifs. Les souches hybrides rouge et blanche d'O. Niloticus ont été développées après une grande sélection artificielle.
Un autre exemple très courant est observé chez les mousquetons: l'hétérozygote pour les allèles de couleur a une fleur rose contrairement au phénotype de couleur rouge et blanc qui apparaîtra si la combinaison de l'allèle est homozygote.
La couleur phénotypique rose semi-dominante apparaîtra si elle est homozygote dominante ou homozygote récessive. Il est également certain que l'un des allèles est dominant et que l'autre est récessif ne s'appliquera pas si un grand nombre d'allèles est présent.
C. Co-dominant:
Parfois, les deux allèles d'un même locus sont co-dominants, les deux contribuant de manière égale au caractère phénotypique des hétérozygotes. Ce modèle d'héritage d'un seul allèle est difficile à distinguer. Pour les éleveurs, l'accouplement doit avoir lieu entre deux homozygotes.
D. Epistasis chez le poisson:
L'épistasie est définie comme la situation qui se produit lorsqu'un allèle d'un gène affecte l'expression des allèles à un autre emplacement / locus sur le même génome. Le phénomène d'épistasie se rencontre couramment chez les animaux et les plantes.
Il existe une interaction fonctionnelle entre différents gènes lorsque l'allèle ou le génotype du locus «masque» ou inhibe l'expression d'un non-allèle ou d'un génotype au niveau d'un locus distinct. L'exemple de ce phénomène se trouve dans les poissons, O. niloticus et Cyprinus carpio.
Chez O. niloticus, les écailles ont un blanc opalescent désigné comme couleur et sont contrôlées épistatiquement au niveau de deux loci. À un locus, l’allèle de type sauvage est désigné par W, tandis que l’autre est Z et l’allèle de type perle est P et à un autre locus, il est L. La couleur perle apparaîtra chez les poissons qui, lors d’un assortiment indépendant, obtiennent des allèles P et L à locus.
Cela n'implique pas le type standard de dominance car l'allèle de type perle peut être exprimé à l'état hétérozygote. C'est simplement la présence d'allèles de type perle au niveau des deux loci qui confère le phénotype perle à son porteur. Au deuxième locus, l'allèle de type sauvage est désigné par Z et l'allèle de type perle par L.
Si la combinaison est WPZL, ce sera perle. Si la combinaison est WPLL, ce sera également une perle. Si la combinaison est en PPZL, ce sera également une perle et si la combinaison est une PPLL, ce sera également une perle. Le rapport entre le phénotype sauvage et celui de la perle sera de 7 (sauvage): 9 (la perle).
C'est simplement la présence des allèles P et L de type phénotype perlé aux deux loci qui expriment le phénotype perle chez les poissons. WWZZ (type sauvage) et PPLL (perle). Le type de perle si les individus doubles hétérozygotes F 1 suivants sont croisés (WPZL croisé en WPZL) (Fig. 40.3).

Un autre exemple d'interaction épistatique est également observé dans le motif d'écailles de la carpe commune, Cyprinus carpio. L'accouplement se fait entre les poissons, qui sont doubles hétérozygotes. L'avantage de ces techniques a également été utilisé en aquaculture.
Certains caractères quantitatifs d’importance commerciale souhaitée ont été produits chez certains poissons, tels que Cyprinus carpio, qui est cultivé dans le monde entier. Cyprinus carpio est le poisson d'élevage le plus important au monde. Il a une grande valeur commerciale. Cyprinus carpio, une carpe majeure exotique, est également bien adoptée dans les eaux indiennes. Il est maintenant élevé avec succès par la reproduction induite dans le monde entier.
Il a trois variétés principales d’importance commerciale comme sous:
(1) Cyprinus Carpio (Communis):
Le nom commun est la carpe écaille. Le corps est recouvert de petites écailles régulièrement disposées. Le motif est de type sauvage. L'allèle de type sauvage est représenté par W tandis que l'allèle variant est désigné par le mot S. À un autre locus, l'allèle de type sauvage est Z et l'allèle variant est N.
L'apparition d'allèles Z modifie le modèle d'échelles chez les hétérozygotes, mais est mortelle à l'état homozygote (ZZ). Ce lieu est épistatique par rapport au W / S et modifie le schéma des écailles, les hétérozygotes (ZN) survivent et les NN homozygotes ont des écailles de type sauvage.
(2) Cyprinus Carpio (Specularis):
Il est communément appelé carpe miroir. Le corps est couvert de grandes écailles jaunes inégalement brillantes, normalement limitées à la ligne latérale, ce qui laisse une grande partie du corps sans écailles.
(3) Cyprinus Carpio (Nudus):
Le nom commun est la carpe en cuir. Le corps est pourvu d'une rangée d'écailles quelque peu dégénérées le long de la base de la nageoire dorsale. Le reste du corps est dépourvu d'échelle. Ainsi, Cyprinus carpio (Nudus) présente un schéma à échelle réduite.
Ces poissons ont une grande valeur sur le marché européen alors que Cyprinus carpio (Communis) ayant un motif de type sauvage est préféré dans les pays asiatiques. Les pisciculteurs, en fonction des besoins du marché, peuvent désormais les produire en utilisant l’utilisation de la combinaison génotypique, son schéma héréditaire et en appliquant les techniques décrites ci-dessus.
Si le croisement est fait entre doubles hétérozygotes, le résultat est décrit comme suit (Fig. 40.4):

E. Une autre situation se présentera si des allèles à deux caractères sont présents sur deux loci mais qu’ils ne sont pas liés ou qu’ils n’ont pas d’influence réciproque:
Dans certains poissons, deux caractères importants ont une valeur commerciale, mais ils sont contrôlés par deux loci et ces loci ne sont pas liés entre eux. Le meilleur exemple de ceci est Lebistes reticulates, un guppy. Les caractères phénotypiques sont la couleur d'or et la courbure de la colonne vertébrale.
Les deux allèles pour la couleur sont W dominante pour sauvage (couleur noire) et G pour la couleur dorée, dont le caractère est récessif. Pour la courbure du poisson, le récessif est représenté par C et le sauvage dominant est représenté par Z.
Si deux hétérozygotes sont croisés. Le résultat serait le suivant (Fig. 40.5):

Ceci indique clairement que deux allèles de gènes présents aux deux loci n'ont aucune influence l'un sur l'autre. L'éleveur peut tirer parti de ce caractère et élever les poissons de manière à ce que seuls les poissons à la couleur de corps dorée avec une colonne vertébrale normale apparaissent dans leurs écloseries.
F. Loci non liés:
Dans les guppys (Les Lebistes réticulent), il y a deux caractères, l'un est de couleur or et l'autre, de courbure de la colonne vertébrale. Ils sont situés et contrôlés indépendamment dans des lieux distincts. Au premier locus, l'allèle dominant de type sauvage (W) produit une coloration grise, tandis que l'allèle récessif (G) produit une coloration dorée chez les poissons homozygotes (GG).
En raison de la domination complète de l'allèle W, le poisson hétérozygote (WG) est gris. Au deuxième locus, une variante récessive (C) provoque la courbure de la colonne vertébrale chez les homozygotes (CC). Le poisson dominant a un poisson à épine normale.
L'héritage quantitatif a également été remarqué chez les poissons. L'avantage des caractères quantitatifs a été utilisé en aquaculture. En général, la couleur de la variété sauvage de Cyprinus carpio est grise, mais dans la population, certains poissons présentent une couleur dorée. La couleur dorée phénotypique est donc un trait qualitatif.
Ces poissons de couleur dorée sont appelés comme variantes. Le variant est un allèle ou un phénotype qui diffère du type standard ou du type sauvage mais qui n'est ni délétère ni anormal. La couleur dorée du phénotype qui diffère de la couleur grise de type sauvage standard est due aux mutations génotypiques des allèles que le poisson obtient par héritage, mais le mutant n’est ni délétère ni anormal, et par conséquent il survit. La variante en or a une valeur marchande élevée.